Broadly, research in the Branch lab aims to understand the evolution of communication signals. Despite having an arguably small set of concerns – to survive and reproduce – animals send and receive information using an impressive range of signals across sensory modalities. Related, we are also interested in the interplay between signaling, sexual selection, and cognitive adaptation. Along with talented collaborators, we aim to understand how and if advanced cognitive abilities, shown to be critical for survival, are conveyed via secondary sexual traits, like male song and variation in plumage. To date, the evidence for secondary sexual traits that provide information about cognitive prowess is equivocal and scant. Part of the Branch lab’s interests are establishing when and how cognitive abilities may be important for mate choice via sexual selection.




Cognitive adaptation
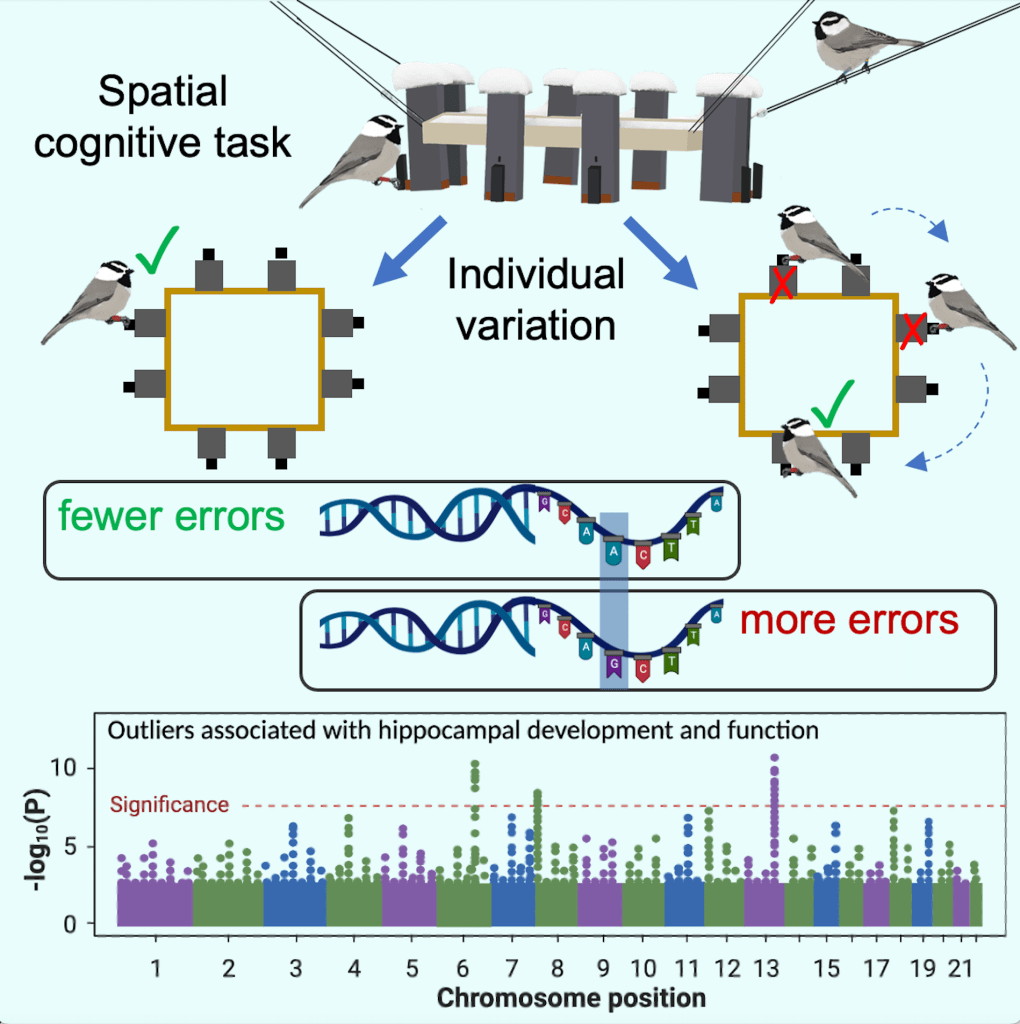
At our long-term field site in the Sierra Nevada mountains (collaboration with Dr. Vladimir Pravosudov at the University of Nevada, Reno), we study spatial cognitive abilities in free-living mountain chickadees (Poecile gambeli). Using innovative ‘smart’ feeder arrays we are able to program individuals to a single feeder within the array and assess their learning and memory performance (Croston et al. 2016, 2017; Tello-Ramos et al. 2018, Animal Behaviour). We assess cognitive ability along an elevation gradient, where selection pressures on overwinter survival increase with elevation (Branch et al. 2019; Benedict et al. 2020, Behavioral Ecology and Sociobiology). Mountain chickadees are nonmigratory, food-caching birds that store thousands of food items throughout their territories during times when food is abundant, to consume later when food is scarce. These birds use their specialized spatial memory abilities to recover their food stores and survive the harsh montane winters. Our research shows that birds inhabiting higher, harsher elevations perform significantly better on our spatial task compared to their lower elevation counterparts.

Recently, we’ve shown that selection is acting on the cognitive abilities of these small birds. Individuals that perform better on our spatial cognitive task are more likely to survive their first winter and into the breeding season (Sonnenberg et al. 2019, Current Biology). We’ve also shown that females increase their reproductive investment when their social mate has better spatial cognition (Branch et al. 2019, Ecology Letters). Furthermore, in collaboration with Drs. Georgy Semenov and Scott Taylor at the University of Colorado, Boulder, we’ve shown that variation in spatial cognition is associated with differences in the genome, known to be related to neuron growth and development (Branch et al. 2022, Current Biology).
Mate preference and sexual selection
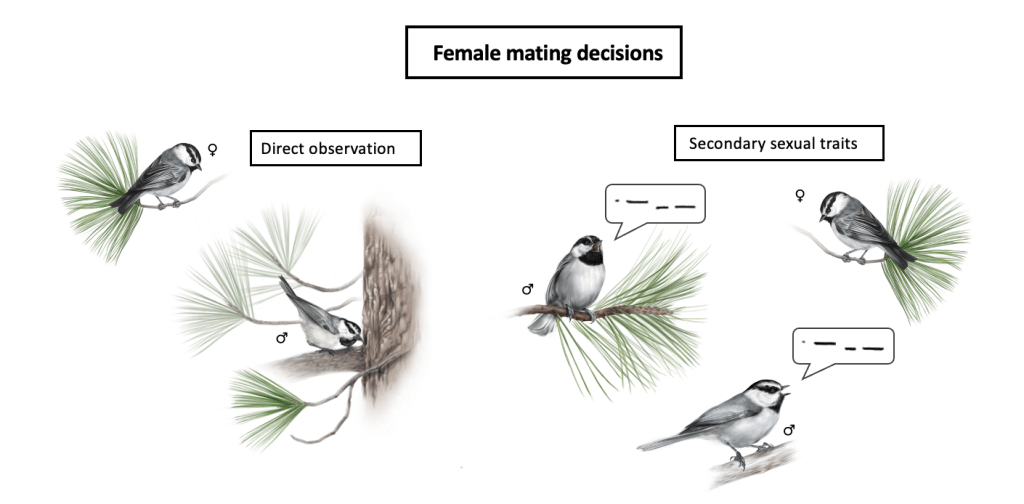
While these birds inhabit a continuous elevation gradient and exhibit some level of historical gene flow (Branch et al. 2017), in 10 years we’ve only documented 2 birds hatching at low elevation that bred at high. This begs the question of whether or not females play a role in the separation we see between high and low elevations – Do females prefer males from their respective elevation? If so, how can they discriminate among high and low elevation males? Or do they prefer males with better cognitive abilities regardless of elevation?
We’ve shown that they can discriminate among high and low elevation males (Branch et al. 2015, Animal Behaviour), and that song (Branch & Pravosudov 2015, Royal Society Open Science; 2020, Behavioral Ecology and Sociobiology) and plumage (Lane et al. in prep) vary between high and low elevation males. Currently, we are working to assess whether females choose extra-pair mates based on their cognitive abilities and whether or not that information is conveyed via secondary sexual traits.
Drivers of complex vocal communication
North American chickadees provide an interesting example of complex vocal communication among Passerines (or songbirds), as they produce relatively simple, whistle songs and complex, syntax following calls. Among temperate songbirds, song is learned by local males and typically thought to be produced primarily in the spring by males to attract females and defend territories from conspecifics. While calls are thought to be innate and are used all year by both sexes.
Example vocal types from mountain chickadees, recorded in the Sierra Nevada


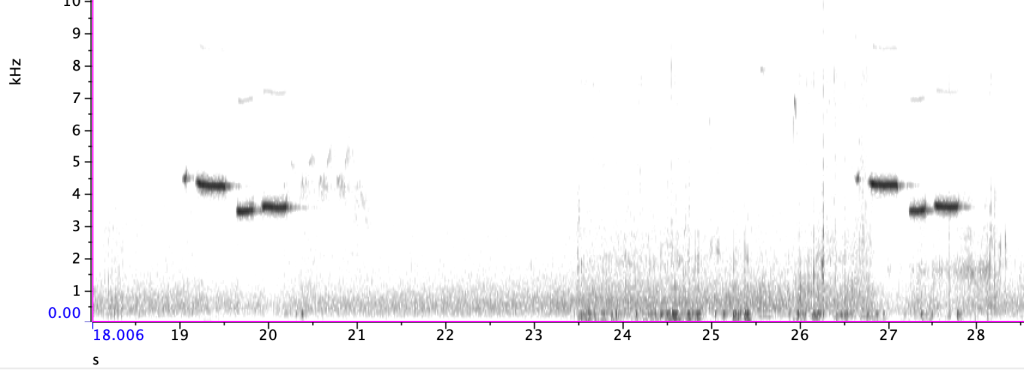
Song is species specific and often thought of as melodic and complex (of course beauty lies in tympanic membrane of the beholder), while calls, and more specifically alarm calls, seem to have converged on a general, highly transmissible structure. And of course, chickadees break these ‘rules’. In addition, chickadees produce additional call types, including seet and gargle calls. Their highly complex vocal system combined with their life history and the previous research of many scientists, provides a prime opportunity to investigate the ecological and environmental drivers of complex vocal communication.
Image credit: Scott Taylor
